Son proliferaciones indiscriminadas de macrófagos o DC. El histiocitoma cutáneo canino es una neoplasia benigna de la epidermis con origen en células de Langerhans.
Los histocitomas son comunes en perros, pero raros en cabras y bovinos. No son malignos, ambas se presentan con lesiones de piel o en tejido subcutáneo. La histiocitosis cutánea no tiene predisposición de raza, se presenta como nódulos no dolorosos aislados o múltiples en la piel o en el tejido subcutáneo.
lunes, 17 de octubre de 2011
Otras células presentadroas
MACRÓFAGOS
Procesan y presentan a linfocitos T sensibilizados, no pueden activar linfocitos vírgenes. Son semi profesionales.
LINFOCITOS B
Pueden procesar grandes cantidades de ag, en asociación con MHC II pueden ingerir y procesar ags antes de presentarlos a los LT sensibilizados.
OTRAS
células “no profesionales” como neutrófilos, eosinófilos, linfocitos T, fibroblastos, células NK, células de músculo liso, astrocitos, células de microglia, y células epiteliales del timo y corneales. Los fibroblastos pueden ser eficaces presentadores dentro de órganos linfoides. Las células del endotelio vascular también pueden captar ags, sintetizar IL1 y por influencia del IFN-y expresar MHC clase II. Incluso los queratocinocitos pueden sintetizar IL-1 y expresar MHC II. En cerdos, hay una población de linfocitos T y/8 que pueden comportarse como presentadores profesionales.
Procesan y presentan a linfocitos T sensibilizados, no pueden activar linfocitos vírgenes. Son semi profesionales.
LINFOCITOS B
Pueden procesar grandes cantidades de ag, en asociación con MHC II pueden ingerir y procesar ags antes de presentarlos a los LT sensibilizados.
OTRAS
células “no profesionales” como neutrófilos, eosinófilos, linfocitos T, fibroblastos, células NK, células de músculo liso, astrocitos, células de microglia, y células epiteliales del timo y corneales. Los fibroblastos pueden ser eficaces presentadores dentro de órganos linfoides. Las células del endotelio vascular también pueden captar ags, sintetizar IL1 y por influencia del IFN-y expresar MHC clase II. Incluso los queratocinocitos pueden sintetizar IL-1 y expresar MHC II. En cerdos, hay una población de linfocitos T y/8 que pueden comportarse como presentadores profesionales.
Células dendríticas en animales domésticos
- Equinos: CMHII, CD11+, EqWC1+, EqWC2+.
- Bovinos: CD80, CD86, CD40, moleculas restringidas a bovinos.
Tienen dos subpoblaciones; una sintetiza mas IL2 y la otra mas IL1 y IL10.- Ovinos:CMH II+, CD11e+ y CD13-.
- Cerdos:
- MDC: CD172a+, CD11R1+,CD1+,CD80+/CDb6+/-.
- PDC: CD172a+, CD4+, CD1+ CD80/86+/-.
Ambos secretan IL10 e IL2.


- Perros: CMH II+, CD34+, CD14-.
- Felinos: langerhans son CD18+, CMHII+, CD1a y CD4.
Procesamiento de antígenos endógenos
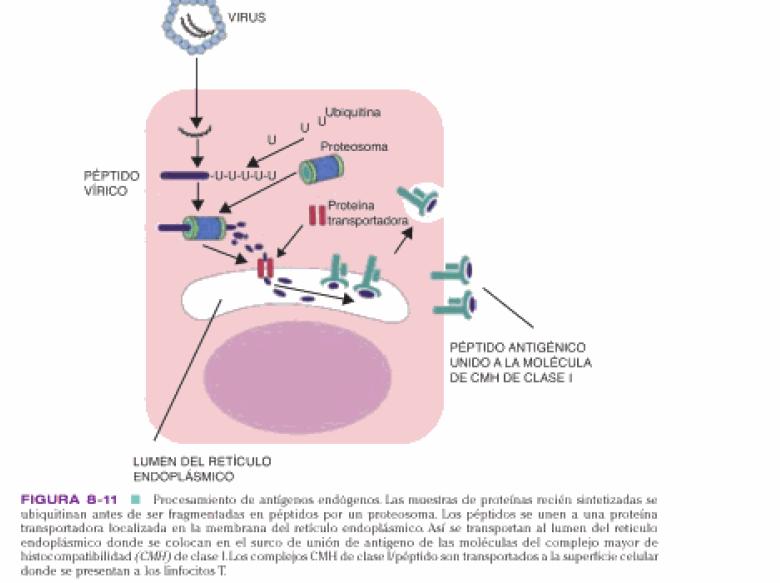
Los LT tienen como una función detectar y destruir células que producen proteínas anormales o extrañas, como los virus. Para controlar la infección, los LT solo responden a células que expresen el CMH 1ª. La cadena a del CMH de clase 1ª se pliega de modo que forma un surco de unión al ag en su superficie externa. Debido a que el surco de unión esta cerrado, las moléculas del MHC solo pueden unirse a 9 aa en su parte media.
Las células vivas fragmentan y reciclan proteínas constantemente, por consiguiente se eliminan las proteínas anormales, los péptidos reguladores no se acumulan y los aa quedan disponibles para otros fines.
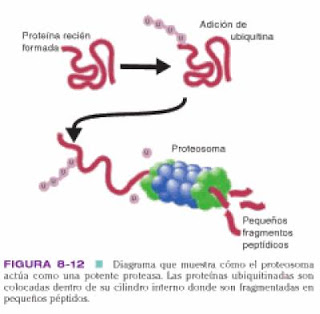
 Primero, las lisinas a reciclar se fijan a la ubiquitina, generalmente se agrega una cadena de 4 moléculas de ubiquitina a cada proteína diana, las proteínas ubiquitinadas son destruidas. Las cadenas son reconocidas por el proteosoma (complejo tubular molecular grande que tienen actividad proteasa y regulan que proteinas se destruyen y cuales no), solo las proteínas ubiquitinadas pueden entrar a los anillos externos; desdoblan la proteína, liberan la ubiquitina y pasan la cadena pepetidica al interior donde se divide en fragmentos de 8 y 15 aa. La actividad de los proteosomas esta regulada por citoquinas, como IFN-y y caspasas. Con el tiempo, los péptidos se protegen de la degradación por su unión a proteínas transportadoras. Se han identificado dos; TAP1 Y TAP2, codificadas por genes localizados dentro del CMH. TAP 1 y 2 seleccionan los péptidos y los transportan desde el citoplasma a los endosomas, aquí los péptidos son atacados por una aminopeptidasa que los acorta en un aa cada vez hasta que se degradan, a no ser que un intermediario, un péptidos de 9 aa encaje con gran precisión en el surco de unión de una molécula de clase I del CMH libre. En este caso, la degradación finaliza y los complejos péptidos-CMH se trasportan a la superficie celular, vía el aparato de Golgi. Una célula puede expresar 104 complejos péptidos-CMH y se necesitan acerca de 200 moléculas de clase I cargadas con el mismo péptidos vírico para activar a un linfocito T citotóxico.
Primero, las lisinas a reciclar se fijan a la ubiquitina, generalmente se agrega una cadena de 4 moléculas de ubiquitina a cada proteína diana, las proteínas ubiquitinadas son destruidas. Las cadenas son reconocidas por el proteosoma (complejo tubular molecular grande que tienen actividad proteasa y regulan que proteinas se destruyen y cuales no), solo las proteínas ubiquitinadas pueden entrar a los anillos externos; desdoblan la proteína, liberan la ubiquitina y pasan la cadena pepetidica al interior donde se divide en fragmentos de 8 y 15 aa. La actividad de los proteosomas esta regulada por citoquinas, como IFN-y y caspasas. Con el tiempo, los péptidos se protegen de la degradación por su unión a proteínas transportadoras. Se han identificado dos; TAP1 Y TAP2, codificadas por genes localizados dentro del CMH. TAP 1 y 2 seleccionan los péptidos y los transportan desde el citoplasma a los endosomas, aquí los péptidos son atacados por una aminopeptidasa que los acorta en un aa cada vez hasta que se degradan, a no ser que un intermediario, un péptidos de 9 aa encaje con gran precisión en el surco de unión de una molécula de clase I del CMH libre. En este caso, la degradación finaliza y los complejos péptidos-CMH se trasportan a la superficie celular, vía el aparato de Golgi. Una célula puede expresar 104 complejos péptidos-CMH y se necesitan acerca de 200 moléculas de clase I cargadas con el mismo péptidos vírico para activar a un linfocito T citotóxico.

Las células vivas fragmentan y reciclan proteínas constantemente, por consiguiente se eliminan las proteínas anormales, los péptidos reguladores no se acumulan y los aa quedan disponibles para otros fines.

PRESENTACIÓN CRUZADA En algunos casos, los ag exógenos pueden entrar al citoplasma y ser procesados y presentados a la molécula CMH 1. Así, en las células presentadoras de ag, los antígenos víricos que han entrado por endocitosis podrían no ser degradados en los lisosomas, sino transportados al citoplasma y se procesarían como ag endógenos. Estos antígenos se quedarían asociados a CMH I y serian reconocidos por los L T citotóxicos.
Procesamiento de antígenos exógenos
La presentacion de ags exogenos esta controlada por las moleculas del CMH clase II. Los L Th reconocen y responden a los fragmentos de ags extraños solo si estan unidos al CMH II.
- El ag se fagocita e incorpora al fagosoma, el cual se fusiona con los lisosomas que tienen proteasas. Estos descomponen los péptidos en varios fragmentos.
- Estos endosomas se fusionan con los del MHCII recién sintetizadas.
- Las cadenas recién sintetizadas del CHMII se transportan al retículo endoplasmático donde se ensamblan en un complejo con un péptidos llamado la cadena invariante (Ii).
- Este complejo se traslada a un endosoma, donde se digieren las moleculas de la Ii, dejando un pequeño péptidos; “péptidos Ii asociado a clase II (CLIP), que permanece asociado a la molecula CMH.
- El CLIP queda en en el sitio antigénico de la molecula del CMH. Cuando los lisosomas que contienen ag se fusionan con los endosomas que tienen CMH, los péptidos antigénicos se cambian por la cadena CLIP.
- EL surco puede albergar de 12-14 aa. Las cadenas CLIP evitan que los endosomas que contienen moleculas CMHII se transporten prematuramente a la superficie celular.
- Una vez unido el ag con el CMH; las vesiculas fusionadas se transportan a la superficie celular, presentando el complejo péptido-CMH.

Células Dendríticas
Tienen 3 funciones:
ORIGEN
Derivan de la célula madre de la médula osea, inmaduras forman redes en todos los tejidos, excepto en cerebro, ojo y testículos.

ESTRUCTURA
Típicamente, pequeño cuerpo celular con muchas dendritas.
Celulas de Langerhans.

MDC vida de 18 meses, estan en epidermis, atrapan y procesan ags que penetran la piel. Influyen en respuestas inmunes cutaneas, como la hipersinsibilidad retardada y dermatitis alergica por contacto. Tienen granulos de Birbeck.
Hay una población de MDC en el timo que reconoce LT autorreactivos e inducir tolerancia en los inmaduros. Pueden expresar CD95L (CD178) para estimular apoptosis de linfocitos T. Su producción esta mediada por IL3.
Los monocitos sanguineos son los precursores de los Mcfs y dc mieloides, en que se conviertan depende de las citoquinas y celulas con las que se tope el monocito mientras madura.
Las MDC recién generadas tienen como papel principal capturar microorganismos invasores ( centinelas ), por su corta vida son “desechables”; si no hallan ag mueren, si si; maduran y se especializan. Tienen muchos receptores que las ayudan a madurar.
Su maduración ocurre en respuesta a alarmitas, como IL-1 y TNF-a y PAMPs. Los tejidos dañados liberan heparan sulfato que se une al TLR4 y activa DC, así también la descomposición de los ácidos nucleicos genera ácido úrico, otro activador de DC.
La HMGB1 es uno de los activadores mas importantes, induce la migración de estas células al lugar de inflamación e invasión microbiana. Las DC inmaduras son atraidas al sitio por las quimioquinas y defensinas.
Las DC inmaduras capturan por fagocitosis, pinocitosis y receptores de superficie pueden capturar cuerpos apóptoticos. Distinguen entre material propio de desecho y agentes externos, por medio de los TLR que se activen, de este modo se asegura tener una respuesta inmune adecuada: el material que no activa TLR no se procesa.
El ph de los fagosomas de DC y LB es alcalino, esto para que el cisteína aspartil proteasas son inhibidoras en este ph el ag se conserva para la reaccion con el CMH.
Celulas dendríticas maduras.
Las DC inmaduras que capturan un ag, lo llevan a lugares para que los LT lo reconozcan. Estas DC son atraidas a organos linfoides por la CCL20 ( el daño tisular puede hacerlo también ) y una vez ahí, maduran.
Las maduras secretan CCL22 para atraer LT, y una vez cerca de ella los envuelve en las dendrítas para que estos busquen restos de ags.
Durante su maduración, las moleculas de CMH se transportan al exterior y se incrementa la superficie celular de moleculas coestimuladoras.


CÉLULAS DENDRÍTICAS FOLICULARES.
Localizadas en folículos linfoides, estimulan activación de linfocito B.
Presentan de dos modos ags a LB: en un animal que no ha sido expuesto al ag, la presentación es un proceso pasivo, solo presentan una superficie para que este se presente. En un animal previamente expuesto al ag, los abs se combinan con el ag para formar complejos ag-sb. Las DC captan estos complejos y los esparcen en estructuras denominadas exosomas. Los LB captan estos exosomas, y después de procesar el ag lo llevan al LT sensibles al ag.

- Centinelas para activar defensas innatas.
- Procesamiento de antígenos exógenos para la RI adquirida.
- Regular ambas formas de respuesta inmune
ORIGEN
Derivan de la célula madre de la médula osea, inmaduras forman redes en todos los tejidos, excepto en cerebro, ojo y testículos.

ESTRUCTURA
Típicamente, pequeño cuerpo celular con muchas dendritas.

- DC mieloides (MDC)
- DC plasmocitoides (PDC)
Celulas de Langerhans.

MDC vida de 18 meses, estan en epidermis, atrapan y procesan ags que penetran la piel. Influyen en respuestas inmunes cutaneas, como la hipersinsibilidad retardada y dermatitis alergica por contacto. Tienen granulos de Birbeck.
Hay una población de MDC en el timo que reconoce LT autorreactivos e inducir tolerancia en los inmaduros. Pueden expresar CD95L (CD178) para estimular apoptosis de linfocitos T. Su producción esta mediada por IL3.
Los monocitos sanguineos son los precursores de los Mcfs y dc mieloides, en que se conviertan depende de las citoquinas y celulas con las que se tope el monocito mientras madura.
- Dc mieloides: cerdos, bovidos, caballos, pollos y perros
- C, langerhans: cerdos, bovidos, caballos, perros, gatos
- DC plasmacitoides: cerdos.
Las MDC recién generadas tienen como papel principal capturar microorganismos invasores ( centinelas ), por su corta vida son “desechables”; si no hallan ag mueren, si si; maduran y se especializan. Tienen muchos receptores que las ayudan a madurar.
Su maduración ocurre en respuesta a alarmitas, como IL-1 y TNF-a y PAMPs. Los tejidos dañados liberan heparan sulfato que se une al TLR4 y activa DC, así también la descomposición de los ácidos nucleicos genera ácido úrico, otro activador de DC.
La HMGB1 es uno de los activadores mas importantes, induce la migración de estas células al lugar de inflamación e invasión microbiana. Las DC inmaduras son atraidas al sitio por las quimioquinas y defensinas.
Las DC inmaduras capturan por fagocitosis, pinocitosis y receptores de superficie pueden capturar cuerpos apóptoticos. Distinguen entre material propio de desecho y agentes externos, por medio de los TLR que se activen, de este modo se asegura tener una respuesta inmune adecuada: el material que no activa TLR no se procesa.
El ph de los fagosomas de DC y LB es alcalino, esto para que el cisteína aspartil proteasas son inhibidoras en este ph el ag se conserva para la reaccion con el CMH.
Celulas dendríticas maduras.
Las DC inmaduras que capturan un ag, lo llevan a lugares para que los LT lo reconozcan. Estas DC son atraidas a organos linfoides por la CCL20 ( el daño tisular puede hacerlo también ) y una vez ahí, maduran.
Las maduras secretan CCL22 para atraer LT, y una vez cerca de ella los envuelve en las dendrítas para que estos busquen restos de ags.
Durante su maduración, las moleculas de CMH se transportan al exterior y se incrementa la superficie celular de moleculas coestimuladoras.

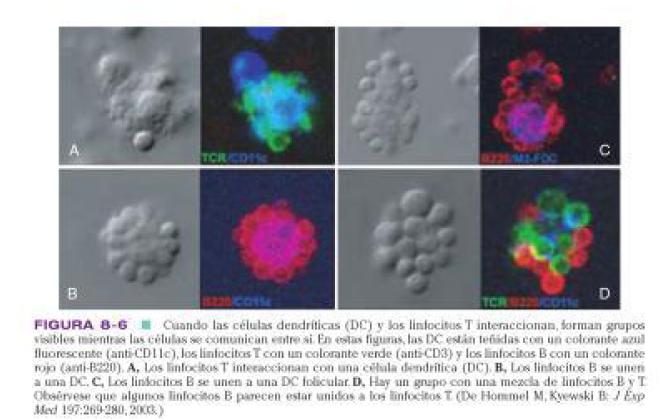
CÉLULAS DENDRÍTICAS FOLICULARES.
Localizadas en folículos linfoides, estimulan activación de linfocito B.
Presentan de dos modos ags a LB: en un animal que no ha sido expuesto al ag, la presentación es un proceso pasivo, solo presentan una superficie para que este se presente. En un animal previamente expuesto al ag, los abs se combinan con el ag para formar complejos ag-sb. Las DC captan estos complejos y los esparcen en estructuras denominadas exosomas. Los LB captan estos exosomas, y después de procesar el ag lo llevan al LT sensibles al ag.
Celulas dendríticas y el procesamiento de antígenos
El organismo tiene un sistema de defensa que reconoce los agentes patógenos sin necesidad de iniciar un proceso inflamatorio ni las molestís que este implica; este es el sistema inmune adquirido. Para que este sistema funcione, se debe captar una muestra de patógeno y ser procesada por las células presentadoras de antígenos.
El procesamiento de antígeno puede seguir aun cuando la amenaza haya terminado, el cuerpo refuerza sus defensas a una segunda exposición.
El procesamiento del antígeno consiste en su fragmentación en péptidos que son capturados y que se unen al complejo mayor de histocompatibilidad. Una vez unidos, son transportadores a la superficie celular. La inmunidad adquirida se desencadena cuando estos péptidos antigénicos unidos al MHC se unen a receptores en los linfocitos.
Los antígenos pueden ser exógenos como las bacterias, o endógenos como los virus producidos por el mismo cuerpo.
Las células que capturan a los exogenos son: DC, macrófagos y linfocitos B. Las mejores son las DC.

El procesamiento de antígeno puede seguir aun cuando la amenaza haya terminado, el cuerpo refuerza sus defensas a una segunda exposición.
El procesamiento del antígeno consiste en su fragmentación en péptidos que son capturados y que se unen al complejo mayor de histocompatibilidad. Una vez unidos, son transportadores a la superficie celular. La inmunidad adquirida se desencadena cuando estos péptidos antigénicos unidos al MHC se unen a receptores en los linfocitos.
Los antígenos pueden ser exógenos como las bacterias, o endógenos como los virus producidos por el mismo cuerpo.
Las células que capturan a los exogenos son: DC, macrófagos y linfocitos B. Las mejores son las DC.

Suscribirse a:
Entradas (Atom)